





-
DNA作为遗传物质,在生命体的生长、发育和繁殖等过程中扮演着核心的角色。由于生命体长期受到内源和外源性遗传毒剂的胁迫,DNA经受着频繁的损伤和突变。为了维持基因组的完整性,生命体进化了一套DNA损伤修复机制——DNA损伤反应(DNA damage response, DDR),该通路包括DNA损伤检验点、信号传导和应激反应三个部分[1]。DNA损伤检验点位于DDR通路的上游,包括hus-1、atm-1、mrt-2和clk-2等,负责损伤信号的识别。而在损伤检验点下游是转导子蛋白(ATM和ATR),把DNA损伤信号进一步级联传递至效应子,诱发细胞应激反应,主要包括细胞周期抑制、细胞凋亡和DNA损伤修复等,其中DNA损伤修复包括非同源末端连接(Nonho-mologous end-joining, NHEJ)、同源重组(Homologous recombination, HR)、错配修复(Mismatch repair, MMR)和碱基切除修复(Base excision repair, BER)等[2]。
目前普遍认为DDR的作用模式是细胞自主性的,即从DNA损伤的感知到下游的应激反应都是以细胞为单位来完成的。但最近越来越多的证据表明DDR也受到胞外信号的调控,具有非细胞自主的调控模式[3-4]。生命体由不同类型的器官、组织和细胞组成,由于其分化程度和功能等的差异,在受到遗传毒剂胁迫时,它们的DDR具有不同的激活程度,这种异质性激活如何影响辐射的生物学效应是目前大家关注的一个重要科学问题。组织/细胞特异性挽救是研究特定生物过程和功能时空特性的重要技术,已经在模式生物上面得到广泛的使用[5]。考虑到DDR激活的细胞/组织依赖性,利用特异性挽救可以建立DDR异质性激活实验体系。
模式生物秀丽隐杆线虫凭借其遗传背景清楚、与人类基因组同源性高、生命周期短、结构简单、突变体资源丰富和易于培养等优势,已经成为科研人员首选的模式生物[6]。启动子是基因表达的开关,具有一定的时空模式,特别是一些组织特性表达启动子,已被广泛用于特定生物学功能的组织特异性挽救体系的构建[7]。在秀丽隐杆线虫中,myo-2基因只在食道球肌肉细胞中表达,其启动子是一个经典的食道球特异表达元件[8-9]。另外,在线虫的体细胞中没有可检测的DDR上游生物活性,HUS-1、ATM-1、ATR-1等DNA蛋白的表达在转录水平被明显抑制[10]。考虑以上因素,在本研究中选择线虫食道球作为目标组织,构建myo-2启动子驱动hus-1基因在食道球特异表达的线虫品系。为了排除体细胞中DDR本底活性的干扰,把其进一步与hus-1突变体杂交,最终建立DDR食道球特异挽救品系。线虫生殖腺位于身体的中部,是生殖细胞发生和受精的部位。精子由精原细胞分化而来,在L4期发育成熟并储存于精囊中。卵母细胞从生殖腺臂的远端到近端依此经历有丝分裂和减数分裂的粗线期、双线期以及终变期,最终发育成熟。这些细胞在各个分裂时期中发生遗传损伤后通常导致胚胎的致死,其致死的程度是评价辐射生物学效应的一个重要指标。在本研究中基于线虫DDR食道球特异挽救品系,以γ射线为辐射源,研究DDR异质性激活对线虫胚胎致死的影响。
-
秀丽隐杆线虫生长培养基(Nematode growth medium, NGM):称取10.2 g琼脂B、1.8 g NaCl、1.5 g蛋白胨用超纯水定容至600 mL,121 °C高压灭菌30 min。培养基温度降至65 °C左右时,在无菌条件下分别加入0.6 mL 1 mol/L的 MgSO4、0.6 mL 1 mol/L的CaCl2、0.6 mL 5 mg/mL的胆固醇、0.6 mL 1%的制霉菌素和15 mL磷酸缓冲溶液(边加边摇匀),然后立即分装到9.0 , 6.0 , 3.5 cm的培养皿中,在超净工作台上放置过夜,待NGM凝固后备用。
M9缓冲溶液:称取1.5 g KH2PO4、3.0 g Na2HPO4、2.5 g NaCl,用超纯水定容至500 mL,121 °C高压灭菌30 min,待温度降至室温时,在无菌条件下加入500 μL 1 mol/L的MgSO4溶液,分装到10个50 mL离心管中,室温保存备用。
-
实验中所用的线虫品系购买自美国国立卫生研究院资助的线虫种质中心(Caenorhabditis genetics center, CGC),涉及到的品系如表1所列。秀丽隐杆线虫在实验室培养条件下以尿嘧啶缺陷型大肠杆菌OP50作为唯一的食物来源,在本研究中避光培养在20 °C生化培养箱中。具体培养方法参照
http://www.wormbook.org 提供的线虫标准实验操作流程。表 1 本实验中所涉及线虫品系
品系名 基因型 描述 N2 野生型 WS2277 hus-1(op241)Ⅰ hus-1基因突变品系 RB2473 cpr-4(ok3413)Ⅴ cpr-4基因突变品系 CB1370 daf-2(e1370)Ⅲ daf-2基因突变品系 BPL1121, BPL1122, BPL1123 Pmyo-2::hus-1::gfp;(op241)Ⅰ hus-1食道球特异挽救线虫品系 BPL1111, BPL1112, BPL1113 Pmyo-2::hus-1::gfp;N2 hus-1食道球特异表达品系 BPL1100 Pmyo-2::gfp;N2 野生型背景下gfp食道球特异表达品系 BPL1101 Pmyo-2::gfp;(op241)Ⅰ hus-1基因突变背景下gfp食道球特异表达品系 BPL1131 Pmyo-2::hus-1::gfp;cpr-4(ok3413)Ⅴ cpr-4基因突变背景下hus-1食道球特异挽救品系 BPL1141 Pmyo-2::hus-1::gfp;daf-2(e1370)Ⅲ daf-2基因突变背景下hus-1食道球特异挽救品系 -
为了使实验过程中所使用的线虫处于同一发育时期以便减少系统误差,我们对线虫进行同步化处理。具体实验过程如下:用M9缓冲液将处于产卵高峰期的线虫从NGM板上洗至1.5 mL离心管中,自然沉降去上清,用M9缓冲液洗三次,加入线虫裂解液(M9缓冲液700 μL、NaClO溶液200 μL、5 mol/L的NaOH溶液100 μL,现配现用)放置在暗处裂解虫体,当70%的虫体裂解时,用离心机6 000 r/min离心1 min,去上清,用M9缓冲液再次清洗三次,然后将虫卵均匀地分散在3 mL M9缓冲液中,过夜孵化后接种在含有OP50的NGM板上,即可得到同步化生长的线虫。
-
将目的序列Pmyo-2::hus-1::gfp(myo-2,食道球特异性启动子,gfp筛选报告基因)插入pPD49.78质粒,然后用显微注射技术将重组质粒打到野生型秀丽隐杆线虫的生殖腺中,得到野生型背景下hus-1在食道球特异性表达的游离型秀丽隐杆线虫株系,其基因型为:Ex[Pmyo-2::hus-1::gfp;N2](转基因操作委托福建上源生物科技有限公司进行)。接着使用γ射线对其进行辐照处理,诱导基因组DNA双链断裂,使游离的质粒通过重组整合到基因组中,筛选三个能够稳定遗传的hus-1食道球特异表达平行转基因品系,根据CGC授权的实验室代码,将其命名为BPL1111、BPL1112和BPL1113[Pmyo-2::hus-1::gfp;N2]。按照此方法构建野生型背景下gfp食道球特异表达品系BPL1100:Pmyo-2::gfp;N2。通过杂交的方法将hus-1食道球特异表达转基因系引入hus-1基因突变背景。在这里我们选择带有绿色荧光蛋白标记的转基因品系BPL1111、BPL1112和BPL1113雄虫作为杂交父本,不带标记的hus-1突变品系WS2277的雌雄同体线虫作为杂交母本,用挑虫针按照3:1的比例挑成虫期的雄虫和L4期的雌雄同体线虫到同一个含有少量OP50的3.5 cm的培养皿中繁殖(增加杂交机率)。当产生后代后,挑一条带绿色荧光标记的F1代雌雄同体线虫单独培养(F1代中带有绿色荧光蛋白标记的即为杂交后代),产生F2代后,再从中挑选24条带绿色荧光蛋白标记的雌雄同体线虫单独培养,产生F3代后,筛选出F3代中均带绿色荧光蛋白标记的F2代线虫,验证其基因型,根据CGC授权的实验室代码,将hus-1食道球特异挽救品系命名为BPL1121、BPL1122和BPL1123[Pmyo-2::hus-1::gfp;(op241)Ⅰ]。按照此方法构建hus-1基因突变背景下gfp食道球特异表达品系BPL1101:Pmyo-2::gfp;(op241)Ⅰ、cpr-4基因突变背景下hus-1食道球特异挽救品系BPL1131:Pmyo-2::hus-1::gfp;cpr-4(ok3413)Ⅴ和daf-2基因突变背景下hus-1食道球特异挽救品系BPL1141:Pmyo-2::hus-1::gfp;daf-2(e1370)Ⅲ。DNA提取方法参照文献[11],在PCR管中加入16 μL RNase-free water和2 μL 10×Ex-Taq bufffer,用挑虫针挑一条线虫放入PCR管中。然后将其放进液氮中1 min,再立即用PCR仪85 °C加热2 min破碎线虫。之后加入1 mg/mL的蛋白酶K溶液2 μL,用PCR仪56 °C加热15 min,95 °C加热10 min,即可得到单条线虫的全基因组DNA。用核酸定量仪Q5000(美国Quawell公司)检测提取的DNA浓度。以单条线虫的全基因组DNA为模板,在PCR管中加入25 μL的2×Hieff PCR Maste Mix(With Dye),2 μL的上游引物(10 μmol/L),2 μL的下游引物(10 μmol/L) ,10 μL的DNA模板,最后加入灭菌后的超纯水至总体积为50 μL。PCR反应程序为:预变性95 °C 2 min,变性94 °C 30 s,退火58.5 °C 30 s,延伸72 °C 50 s,变性-退火-延伸扩增29个循环,终延伸72 °C 2 min。PCR扩增引物如表2所列。扩增产物进行琼脂糖凝胶电泳鉴定,点突变序列送公司测序,并将测序结果上传到NCBI上进行序列比对。
表 2 PCR引物序列
基因 引物 引物序列(5′-3′) hus-1 正向 TGCAGGGAAATAGTGTTCTCA 反向 TCGGCTTCTGGATTTCTAAC daf-2 正向 ATCAATGCGTACTCCTCATC 反向 TGGAAATCTCCGTGACTATC cpr-4 正向 AAAAGACCATCGCAATGAAG 反向 TTCATGATCCTATCAGTCCACG -
将待检测的L3期线虫置于6 cm的NGM板上,利用中国科学院合肥物质科学研究院的Biobeam Cs137辐射源发出的γ射线以3.37 Gy/min的剂量率进行辐照,诱导DNA损伤产生。辐照结束后将线虫转移到新鲜的含有OP50的NGM板上,放置在20 °C培养箱中避光培养。用于后续胚胎致死率的检测。
-
本研究以秀丽隐杆线虫的胚胎致死率作为检测终点,参照文献[12]的实验方法:用挑虫针挑取30条待检测的L4期雌雄同体线虫放在含有OP50的3.5 cm的NGM板上单独培养,在体视镜下记录每条线虫第一颗受精卵排出的时间,在培养箱中产卵24 h后移走母虫,放回培养箱中孵化24 h,然后在培养板上加入40 mmol/L的NaN3溶液30 μL对线虫麻醉5 min。在体视镜下统计出幼虫的数目和未孵化的受精卵的数目,总产卵数等于孵化的幼虫数目和未孵化的受精卵的数目之和,胚胎致死率等于未孵化的受精卵的数目与总产卵数的比值。
-
本实验所有数据的分析均采用GraphPad Prism 6.0软件进行统计学差异分析,均采用平均值±标准差(Mean±Standard Deviation)的形式表示。组间差异分析使用双边t检验的统计学方法,当P值小于0.05时,表示两组数据间具有统计学差异。
-
我们首先检测了γ辐射对DDR食道球挽救品系(BPL1121)胚胎致死的影响。L3期的秀丽隐杆线虫分别暴露于0, 10, 20和40 Gy的γ辐射。结果如图1(a)所示,三个品系的胚胎致死率与辐射剂量呈正相关,BPL1121的胚胎致死率明显高于N2和hus-1基因突变线虫,并且BPL1121在20 Gy时最先达到半数致死量。以20 Gy辐照处理为例,hus-1基因突变线虫的胚胎致死率相对于野生型线虫增加了1.48倍(P<0.01),而BPL1121线虫的胚胎致死率相对于N2增加了3.59倍(P<0.001),相对于hus-1突变体线虫增加了0.85倍(P<0.01)。考虑到稳定转基因线虫过程中,目的基因的整合是随机的,为了排除目的基因序列插入位点的影响,我们检测了20 Gy γ辐照诱导BPL1121的平行品系BPL1122和BPL1123的胚胎致死水平。结果如图1(b)所示,BPL1122和BPL1123的胚胎致死率相对于hus-1突变线虫分别增加了0.78和0.63倍(P<0.01),与BPL1121表现出相同的趋势。另外,为了排除hus-1以及报告基因gfp食道球过表达本身对胚胎致死的影响,我们检测了20 Gy辐照对N2、BPL1111、BPL1112、BPL113、BPL1100和BPL1101的胚胎致死水平,发现阳性对照品系BPL1111、BPL1112、BPL113、BPL1100与N2相比胚胎致死率均没有统计学意义上的差异(P>0.05)[图1(c)和(d)],阴性对照品系BPL1101与hus-1突变品系胚胎致死率没有明显的差异(P>0.05)[图1(e)],表明辐射诱导BPL1121高胚胎致死主要来自DDR的食道球特异性挽救。

图 1 hus-1异位表达增强线虫生殖细胞的辐射敏感性(在线彩图)
-
辐射也能诱导基因组不稳定,在后代中表现出增加的遗传损伤。为了研究DDR食道球挽救线虫中辐射诱导胚胎致死的遗传效应,我们进一步检测了辐照处理BPL1121线虫的F1代和F2代胚胎致死情况。结果如图2所示。经20 Gy γ射线辐照后,与hus-1突变体线虫相比,辐射当代BPL1121线虫的胚胎(P0)致死率由24.72±1.77增加到42.66±1.18(P < 0.01),F1代BPL1121线虫的胚胎致死率由1.55±0.18增加到4.71±0.63(P < 0.01),F2代BPL1121线虫的胚胎致死率恢复到了未辐照时的水平。上述研究结果表明,γ射线辐照DDR食道球挽救线虫BPL1121可能使辐照当代生殖细胞的遗传物质发生了突变,使F1代表现出类似于辐照当代的生物学效应。已有的研究表明,当代生物体暴露于环境胁迫后可以产生跨代遗传效应。关于DNA损伤突变的跨代遗传效应目前主要解释为辐射或其它胁迫引起的基因组不稳定性。现有的报道中,跨代遗传效应大多可以持续3~5代。图2显示,在三个品系中,F1代的胚胎致死率相对于P0代均有明显的降低,F2代已恢复到了未辐照时的水平,说明在我们的实验条件下,基因组不稳定只持续了1代,这可能也是DDR异质性的一个特点,相关详细的机制还有待进一步的研究。
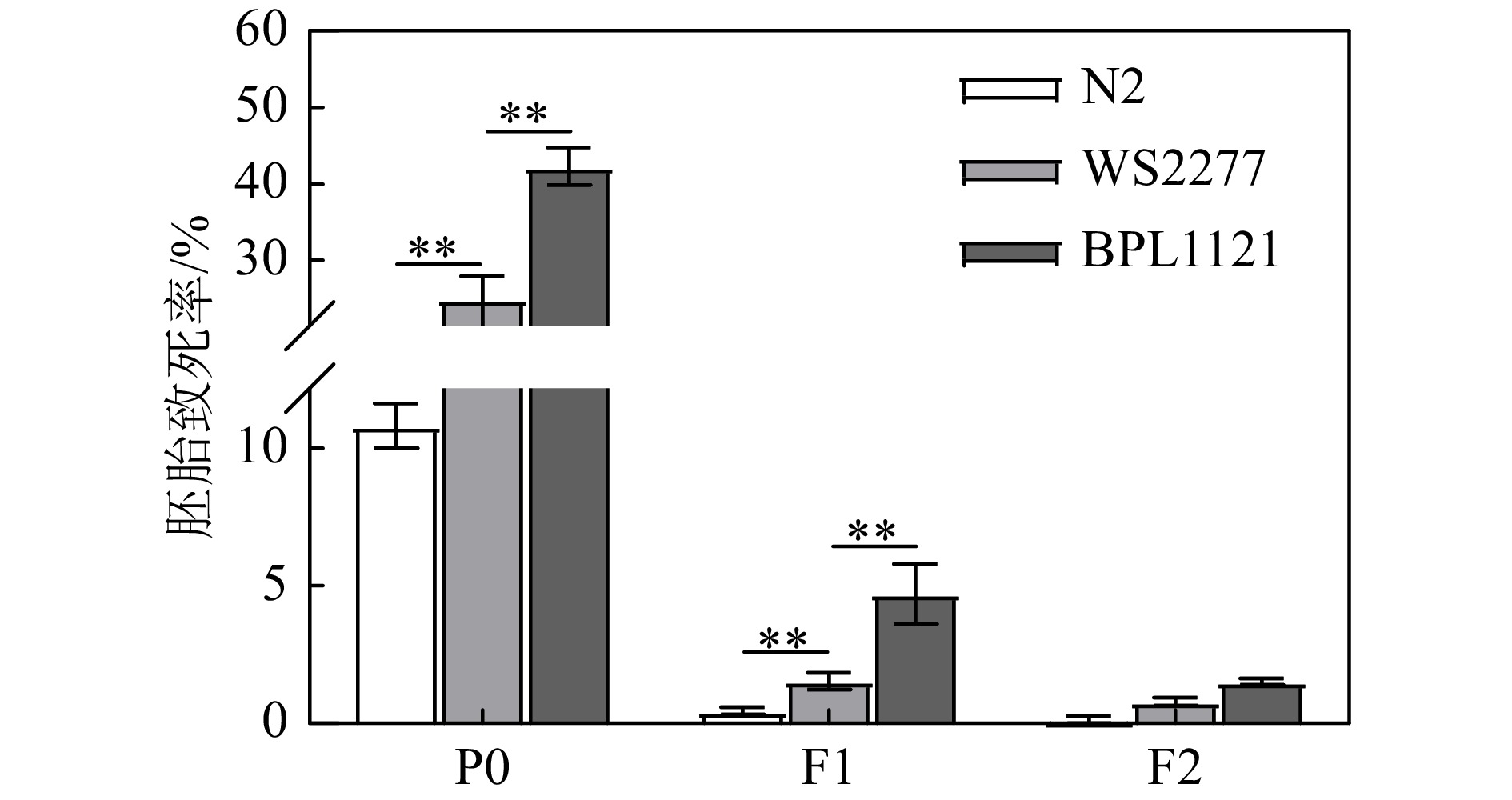
图 2 hus-1食道球挽救对辐射诱导的线虫胚胎致死的遗传效应
-
2017年,清华大学薛定教授课题组发现了线虫中第一个参与辐射旁效应的因子—人类组织蛋白酶B的同源物cpr-4。cpr-4是紫外线或电离γ射线作用于线虫时分泌的,也是条件培养基中导致细胞死亡和未辐照线虫中胚胎致死的主要原因,他们认为cpr-4可能是通过类胰岛素样生长因子受体daf-2来介导辐射旁效应的[13]。基于此,我们选择BPL1121的雄虫作为杂交父本,cpr-4基因突变的线虫品系RB2473作为杂交母本,利用聚合酶链式反应PCR的方法筛选出杂交纯和突变体,将BPL1121构建在cpr-4突变(RB2473)背景下,根据CGC授权的实验室代码将BPL1121引入cpr-4突变背景的线虫品系命名为BPL1131:Pmyo-2::hus-1::gfp;cpr-4(ok3413)Ⅴ。相关线虫品系经20 Gy γ射线辐照处理后,cpr-4突变体线虫的胚胎致死率与N2相比增加了0.63倍(P<0.05),DDR食道球挽救品系BPL1121的胚胎致死率与cpr-4突变体相比增加了1.88倍(P<0.001),与cpr-4突变背景下DDR食道球挽救品系BPL1131相比胚胎致死率没有统计学意义的差异(P>0.05)(图3)。上述研究结果表明,cpr-4没有参与DDR食道球特异挽救对辐射诱导的线虫胚胎致死的影响。
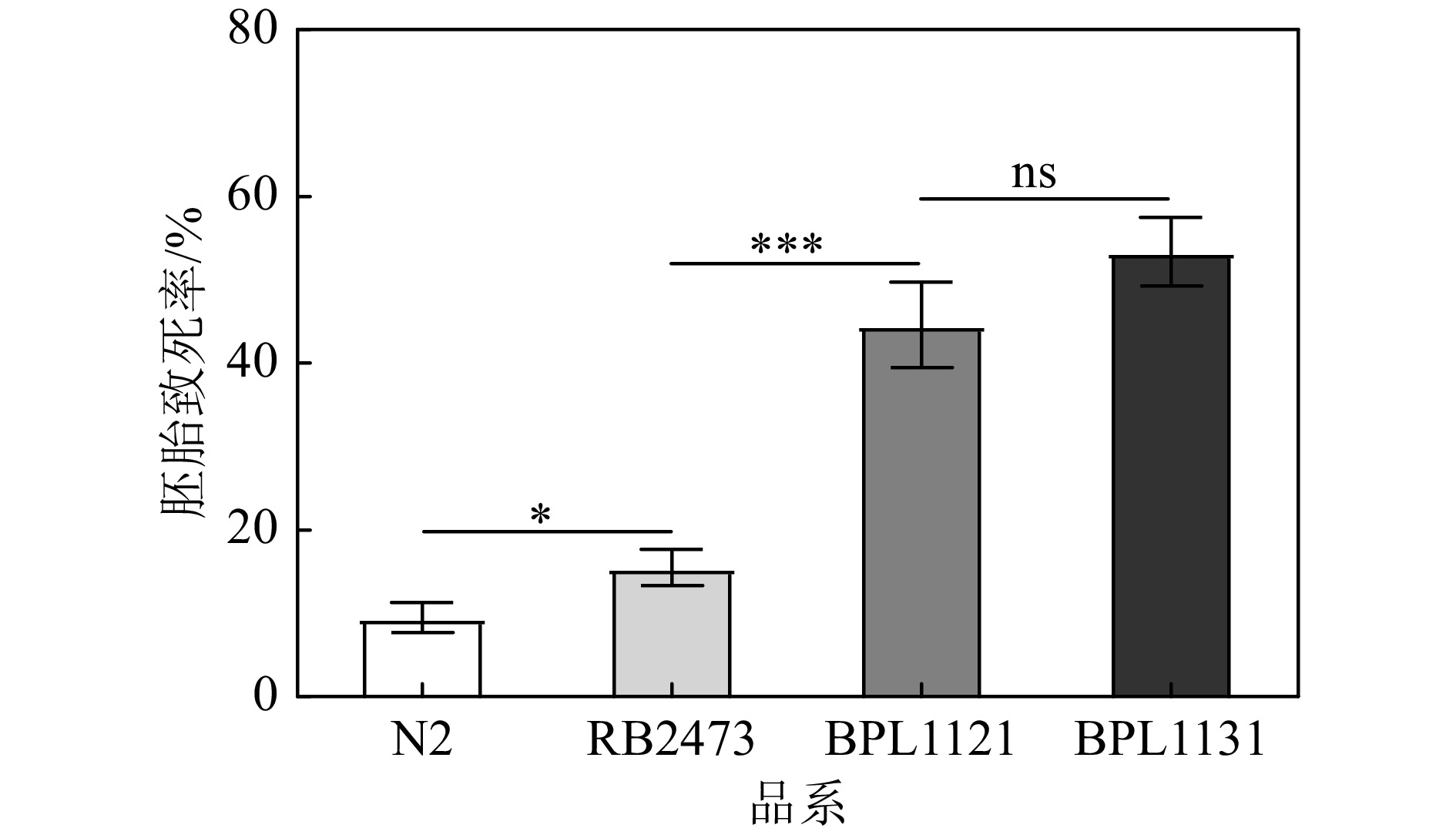
图 3 cpr-4不参与hus-1食道球挽救对线虫生殖细胞辐射敏感性的调控
-
接下来,我们选择BPL1121的雄虫作为杂交父本,daf-2基因突变的线虫品系CB1370作为杂交母本,利用PCR的方法筛选出杂交纯和突变体,将BPL1121构建在daf-2突变(CB1370)背景下,根据CGC授权的实验室代码将BPL1121引入daf-2突变背景的线虫品系命名为BPL1141:Pmyo-2::hus-1::gfp;daf-2(e1370)Ⅲ。相关线虫品系经20 Gy γ射线辐照处理后,daf-2突变体线虫的胚胎致死率与N2相比增加了0.46倍(P<0.05),DDR食道球挽救品系BPL1121的胚胎致死率与daf-2突变体相比增加了2.21倍(P<0.001),与daf-2突变背景下DDR食道球挽救品系BPL1141相比增加了0.40倍(P<0.05)(图4)。上述研究结果表明,daf-2参与了DDR食道球特异挽救对辐射诱导的线虫胚胎致死的影响。

图 4 daf-2参与hus-1食道球挽救对线虫生殖细胞辐射敏感性的调控
-
一个细胞的DNA每天都发生上万次的损伤,包括碱基损伤(Base Damage, BD)、单链断裂(Single-strand breakage, SSB)、链间交联(Interstrand cross-linking, ICL)和DNA双链断裂(Double-strand breakage, DSB)等[14]。为了应对这些损伤,生命体进化了高度复杂的DDR机制。由于DDR在维持基因组稳定中的关键作用,它的许多组分都是重要的肿瘤抑制因子。然而,构成生命体的各种细胞具有不同分化程度和功能,这也就决定了在受到遗传毒剂胁迫时,它们中的DDR具有不同的激活特性,而身体DDR的这种异质性会如何影响遗传毒剂以及辐射的生物学效应,一直是一个有待解决的科学问题。在本研究中,我们建立了一个DDR食道球特异挽救线虫品系来模拟DDR的异质性激活的特点。在这个模型中,我们选择食道球作为DDR的挽救组织,主要是基于以下考虑:1) 秀丽隐杆线虫身体透明,在光学显微镜下可轻易辨别食道球[15];2) 食道球结构简单且表征良好。食道球由95个细胞组成,其位置和形态已经在超微结构水平上进行了表征[16];3) 食道球发育稳健,在其它组织中发育异常的胚胎仍然可以产生分化的食道球[17-18];4) 具有可靠的在食道球特异表达的基因启动子。更重要的是,DDR通路中的CEP-1/p53在线虫食道球中有微量的活性,说明食道球相对于其它的身体组织(生殖腺除外)有相对高的DDR活性[19]。因此,在本研究中,我们选择食道球作为DDR的挽救组织是合适的。
生殖细胞中的DNA损伤不能被有效修复和及时清除,通常会导致胚胎致死。因此,DDR通路对线虫胚胎的发育具有保护作用,它的一些关键基因,如hus-1等的突变能够增加辐射诱导的线虫胚胎致死[20]。在目前的研究中,我们的结果也证明了这一点[图1(a)]。然而令人不解的,是DDR在食道球的特异挽救并没有缓解生殖细胞的辐射损伤,反而加剧了胚胎的致死。与身体细胞不同,线虫的生殖细胞具有完善的DDR通路,在其上游包括两个关键的信号级联:Rad9-Rad1-Hus1(9-1-1)-ATR和MRE11/RAD50/NBS1(MRN)-ATM。在我们构建的DDR挽救品系中,食道球具有较强的9-1-1-ATR活性,但没有明显的MRN-ATM活性。然而在生殖细胞中,9-1-1-ATR通路的功能是缺失的,而MRN-ATM具有正常的功能。因此,我们推测:在DDR挽救品系中,食道球的9-1-1-ATR信号级联远程下调了生殖腺MRN-ATM的功能,降低了辐射损伤的修复效率,进而引起了严重的胚胎致死,前期研究也证明9-1-1-ATR可以远程调控线虫身体其它组织的ATM通路的活性[21]。
CPR-4作为人组织蛋白酶B的同源物,已经被证明介导了线虫的辐射旁效应[13]。为了进一步探究CPR-4在DDR的食道球—生殖腺调控中的角色,我们检测了线虫品系BPL1131的胚胎致死情况,发现cpr-4基因突变并不影响BPL1131线虫的胚胎致死,说明CRP-4并没有参与DDR的食道球—生殖腺调控。线虫中的类胰岛素通路与线虫的生长发育、繁殖、寿命、营养代谢和行为等密切相关,同时也参与了线虫对内外源胁迫的响应,daf-2是该信号通路上的一个关键基因。为了进一步证明类胰岛素途径是否参与DDR的食道球—生殖腺调控,我们检测了线虫BPL1141品系,发现daf-2基因突变明显降低了线虫BPL1141的胚胎致死率,说明线虫食道球9-1-1-ATR通过类胰岛素通路远程负调控了生殖细胞的MRN-ATM,但其具体的作用机制尚待进一步深入研究。
总的来讲,在本研究中,我们利用线虫DDR特异挽救品系,初步明确了DDR异质性对辐射生物效应的影响。该研究对肿瘤的治疗也有一定的借鉴意义,例如,在肿瘤微环境中,除了肿瘤细胞外,还包括周围的成纤维细胞、巨噬细胞、免疫细胞和肿瘤血管等。这些细胞在肿瘤细胞的发生、发展和转移的过程中有重要的作用[22]。众所周知,肿瘤细胞的DDR几乎都存在某种缺陷,但微环境中的其它细胞DDR功能通常是正常的,这表明在肿瘤微环境中DDR的激活也是异质性的。因此,深入研究DDR异质性对辐射效应的影响也能为癌症放疗提供一些新的理论支撑。
Studies on Effect and Its Releated Mechanism of Pharynx-specific Rescue of DDR on Radiation-induced Embryonic Lethality in Caenorhabditis Elegans
-
摘要: DNA损伤反应(DDR)是生命对抗外源和内源性氧化胁迫的主要机制之一。在生命体的不同细胞和组织中DDR具有不同的激活程度,本研究以模式生物秀丽隐杆线虫为实验对象,通过构建DDR通路上游关键基因hus-1的食道球特异表达转基因系,并将其杂交到hus-1突变背景获取DDR食道球特异挽救品系BPL1121、BPL1122和BPL1123,探究DDR异质性激活对辐射生物效应的影响及其内在机制。利用该实验体系检测DDR食道球挽救对γ辐射诱导胚胎致死的影响。与hus-1突变体线虫相比,辐射当代BPL1121线虫的胚胎致死率由24.72±1.77增加到42.66±1.18(P<0.01),这种增强效应延续到了未经过直接辐照的F1子代线虫(P<0.01),但在F2线虫中恢复到本底水平(P>0.05)。线虫中的类胰岛素通路与线虫的生长、发育和繁殖等行为密切相关,daf-2是该信号通路上的一个关键基因。把BPL1121引入到daf-2突变背景,发现DAF-2功能缺失明显抑制了BPL1121线虫的胚胎辐射致死。DDR食道球异质性激活增加了生殖细胞的辐射敏感性,类胰岛素途径参与了食道球DDR对生殖细胞辐射敏感性的远程调控。Abstract: DNA damage response (DDR) is one of the main mechanisms, by which life counters against exogenous and endogenous oxidative stress induced DNA damage. In an organism, the DDR in different cells and tissues has various levels of activation. . Using the model organism Caenorhabditis elegans as experimental objects, we constructed a transgenic line for the pharynx-specific expression of hus-1 gene, upstream of the DDR pathway, and then introduced into hus-1 mutation background, generating DDR pharynx-specific rescue lines BPL1121, BPL1122 and BPL1123 . The experimental system was used to detect the effect of DDR pharynx rescue on the embryos lethality of γ radiation induced and investigate the effects of heterogeneous activation of DDR on radiation biological effects and its underlying mechanisms. Results showed that the embryonic lethality of irradiated BPL1121 worms increased from 24.72 ± 1.77 to 42.66 ± 1.18 (P < 0.01) compared with hus-1 mutant worms, this enhancement continued up to the intact F1 progeny of non-irradiated worms (P < 0.01), although F2 progeny return to background levels (P > 0.05). The insulin-like pathway in nematodes is closely related to the growth, development and reproduction behaviors of nematodes, and daf-2 is a key gene in this signaling pathway. Introducing BPL1121 into the background of daf-2 mutation, it was found that DAF-2 functional loss significantly inhibited the embryonic radiation lethality of BPL1121 worms. The heterogeneous activation of DDR in pharynx increases the radiation sensitivity of germ cells, and the insulin-like pathway participates in the remote regulation of pharyngeal DDR to radio-sensitivity of germ cells.
-
图 1 hus-1异位表达增强线虫生殖细胞的辐射敏感性(在线彩图)
(a) 辐射诱导线虫胚胎致死率的剂量曲线;(b) BPL1121平行品系的胚胎致死率;(c) hus-1食道球表达品系的胚胎致死率;(d) 野生型背景下食道球表达gfp的胚胎致死率;(e) hus-1突变背景下食道球表达gfp的胚胎致死率。**两个品系之间的胚胎致死率差异,P < 0.01;ns两个品系之间的胚胎致死率差异,P > 0.05。
图 3 cpr-4不参与hus-1食道球挽救对线虫生殖细胞辐射敏感性的调控
* 两个品系之间的胚胎致死率差异,P<0.05;*** 两个品系之间的胚胎致死率差异,P<0.001;ns两个品系之间的胚胎致死率差异,P>0.05。
表 1 本实验中所涉及线虫品系
品系名 基因型 描述 N2 野生型 WS2277 hus-1(op241)Ⅰ hus-1基因突变品系 RB2473 cpr-4(ok3413)Ⅴ cpr-4基因突变品系 CB1370 daf-2(e1370)Ⅲ daf-2基因突变品系 BPL1121, BPL1122, BPL1123 Pmyo-2::hus-1::gfp;(op241)Ⅰ hus-1食道球特异挽救线虫品系 BPL1111, BPL1112, BPL1113 Pmyo-2::hus-1::gfp;N2 hus-1食道球特异表达品系 BPL1100 Pmyo-2::gfp;N2 野生型背景下gfp食道球特异表达品系 BPL1101 Pmyo-2::gfp;(op241)Ⅰ hus-1基因突变背景下gfp食道球特异表达品系 BPL1131 Pmyo-2::hus-1::gfp;cpr-4(ok3413)Ⅴ cpr-4基因突变背景下hus-1食道球特异挽救品系 BPL1141 Pmyo-2::hus-1::gfp;daf-2(e1370)Ⅲ daf-2基因突变背景下hus-1食道球特异挽救品系  下载: 导出CSV
下载: 导出CSV
表 2 PCR引物序列
基因 引物 引物序列(5′-3′) hus-1 正向 TGCAGGGAAATAGTGTTCTCA 反向 TCGGCTTCTGGATTTCTAAC daf-2 正向 ATCAATGCGTACTCCTCATC 反向 TGGAAATCTCCGTGACTATC cpr-4 正向 AAAAGACCATCGCAATGAAG 反向 TTCATGATCCTATCAGTCCACG
下载: 导出CSV
-
[1] HOEIJMAKERS J H J. Nature, 2001, 411: 366. doi: 10.1038/35077232 [2] WANG Z, ZHAO J, WANG G, et al. Cancer Research, 2018, 78(22): 6486. doi: 10.1158/0008-5472.Can-18-1814 [3] REINHARDT H C, SCHUMACHER B. Trends in Genetics, 2012, 28(3): 128. doi: 10.1016/j.tig.2011.12.002 [4] OU H L, SCHUMACHER B. Blood, 2018, 131(5): 488. doi: 10.1182/blood-2017-07-746396 [5] CHOE J H, WATCHMAKER P B, SIMIC M S, et al. Science Translational Medicine, 2021, 13(591): eabe7378. doi: 10.1126/scitranslmed.abe7378 [6] GUPTA B P, REZAI P. Micromachines, 2016, 7(7): 127. doi: 10.3390/mi7070123 [7] 苏春明. 线虫神经元特异性表达体系的建立与应用[D]. 武汉: 华中科技大学, 2014. SU Chunming. The Establishment and Application of C. Elegans Neuron-secific Expression System[D]. Wuhan: Huazhong University of Science and Technology, 2014. (in chinese) [8] MILLER D M, ORTIZ I, BERLINER G C, et al. Cell, 1983, 34(2): 477. doi: 10.1016/0092-8674(83)90381-1 [9] ARDIZZI J P, EPSTEIN H F. Journal of Cell Biology, 1987, 105(6): 2763. doi: 10.1083/jcb.105.6.2763 [10] VERMEZOVIC J, STERGIOU L, HENGARTNER M O, et al. Cell Death & Differ, 2012, 19(11): 1847. doi: 10.1038/cdd.2012.69 [11] 王江岭, 张建成, 顾建锋. 植物检疫, 2011, 25(2): 32. WANG Jiangling, ZHANG Jiancheng, GU Jianfeng. Plant Quarantine, 2011, 25(2): 32. (in Chinese) [12] HOLWAY A H, KIM S H, LA VOLPE A, et al. Journal of Cell Biology, 2006, 172(7): 999. doi: 10.1083/jcb.200512136 [13] PENG Y, ZHANG M, ZHENG L, et al. Nature, 2017, 547: 458. doi: 10.1038/nature23284 [14] DE BONT R, VAN LAREBEKE N. Mutagenesis, 2004, 19(3): 169. doi: 10.1093/mutage/geh025 [15] SULSTON J E, SCHIERENBERG E, WHITE J G, et al. Developmental Biology, 1983, 100(1): 64. doi: 10.1016/0012-1606(83)90201-4 [16] ALBERTSON D G, THOMPSON J N. Philosophical Transactions of the Royal Society B, Biological Sciences, 1976, 275(938): 299. doi: 10.1098/rstb.1976.0085 [17] AHNN J, FIRE A. Genetics, 1994, 137(2): 483. doi: 10.1093/genetics/137.2.483 [18] BAUM P D, GARRIGA G. Neuron, 1997, 19(1): 51. doi: 10.1016/S0896-6273(00)80347-5 [19] TANG H Q, CHEN L W, LIU J L, et al. Radiation Research, 2016, 185(4): 402. doi: 10.1667/rr14368.1 [20] 陈亮稳. 秀丽隐杆线虫挥发性信息素介导辐射旁效应和适应性反应偶联过程和机制研究[D]. 合肥: 中国科学技术大学, 2018. CHEN Liangwen. The Interaction of Radiation-induced Bystander Effects and Adaptive Responses Mediated by Volatile Pheromones in Caenorhabditis Elegans[D]. Hefei: University of Science and Technology of China, 2018. (in Chinese) [21] DAI Z Y, ZHANG W J, SHANG M K, et al. International Journal of Molecular Sciences, 2022, 23(14): 7544. doi: 10.3390/ijms23147544 [22] 杨芳, 于雁. 中国肺癌杂志, 2015, 18(1): 48. doi: 10.3779/j.issn.1009-3419.2015.01.08 YANG Fang, YU Yan. Chinese Journal of Lung Cancer, 2015, 18(1): 48. (in Chinese) doi: 10.3779/j.issn.1009-3419.2015.01.08 -
 点击查看大图
点击查看大图

图(4) / 表 (2)
计量
- 文章访问数: 309
- HTML全文浏览量: 80
- PDF下载量: 13
- 被引次数: 0
 甘公网安备 62010202000723号
甘公网安备 62010202000723号